
Overview
Our lab studies how the brain monitors changes in body temperature (via thermosensitive neurons), blood sodium (via sodium-sensitive neurons) and extracellular fluid osmolality (via osmosensory neurons). We aim to define how these neurons convert these physicochemical parameters into electrical signals that can be transmitted synaptically to central homeostatic neurons that regulate body hydration (e.g. to neurons that control thirst and vasopressin secretion) and body temperature (e.g. neurons that regulate non-shivering thermogenesis).
We also study how the brain’s master time-keeping clock (the suprachiasmatic nucleus) regulates hydration status and body temperature across the sleep–wake cycle. Many of these mechanisms are studied in the context of diseases that involve osmo- and thermoregulatory dysfunction; including dehydration, salt‑dependent hypertension, brain trauma/concussion, or exposure to drugs such as MDMA (ecstasy).
Background
Role of the Hypothalamus in Health and Disease
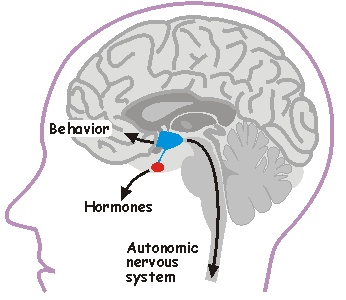
The hypothalamus is a small brain region that regulates many vital body functions such as body temperature, blood pressure, body weight, plasma osmolality, reproduction, sleep, stress responses, and circadian rhythms. It influences behavior, hormone release, and autonomic output.
Systemic Osmoregulation in Health and Disease
Systemic osmoregulation maintains blood osmolality near 300 mosm/kg. Small changes occur after drinking water or consuming salt; larger changes can be dangerous, causing cellular swelling or shrinkage, neurological symptoms, seizures, coma, or death.
Hypothalamic Control of Osmoregulation
Osmoreceptors, sodium detectors, and thermoreceptors in the hypothalamus transduce physiological stimuli into electrical signals that regulate thirst, salt appetite, sympathetic output, and hormone release.
Current Research Topics
Osmosensory Transduction
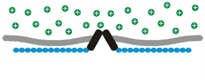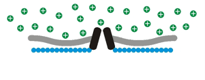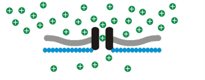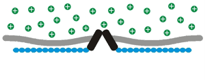
We investigate the cellular and molecular mechanisms by which osmoreceptors detect changes in extracellular osmolality. Our work has shown that mechanosensitive, stretch‑inhibited channels depolarize osmoreceptors under hypertonic conditions. We also study the role of trpv1 and the actin cytoskeleton in osmosensory transduction.
Glia–Neuron Signaling and Osmosensing
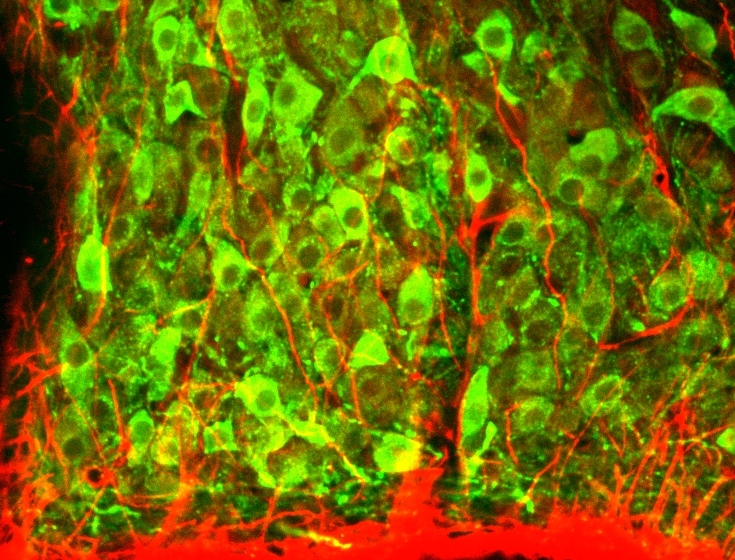
Glial cells release taurine during hypotonicity, activating inhibitory glycine receptors. We study how glia and glia‑derived molecules influence osmosensory neurons, particularly in the supraoptic nucleus where astrocytic processes (red) intercalate between vasopressin and oxytocin neurons (green).
Thermosensory Transduction

TRPV channels are activated by near‑physiological heat. Our discovery of trpv1 expression in supraoptic neurons led to the hypothesis that these neurons are thermosensitive. Ongoing work examines their role in physiological thermosensation.
Synaptic Signals in Osmoregulatory Networks
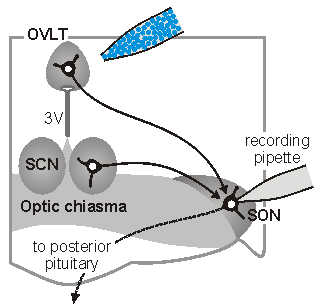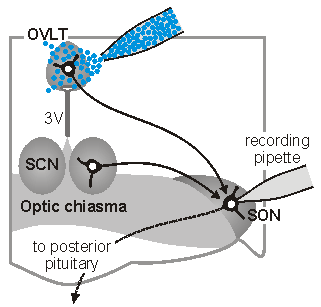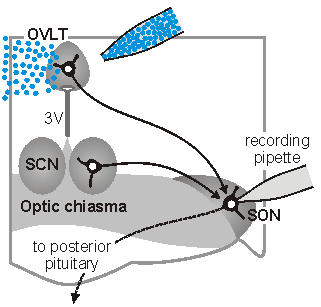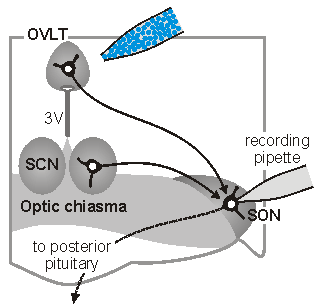
We study how sensory information is transmitted from osmoreceptors and sodium detectors to downstream neurons that regulate behavior, endocrine responses, and autonomic output. We also examine how circadian rhythms modulate synaptic gain in these pathways.
One approach involves stimulating OVLT osmoreceptors while recording synaptic responses in vasopressin neurons of the SON. This work has revealed circadian modulation of osmotic activation. Coverage of this research appears in Scientific American and Science News.
Outcomes
Basic research with clinical relevance
The objective of our research program is to define the neurobiology of hypothalamic circuits that mediate fluid homeostasis. While we focus on fundamental issues related to these networks at the molecular, cellular and systems level, many of our studies are currently performed in the context of important clinical problems including: Salt-dependent hypertension, dehydration, circadian clock disruption, sleep defects, as well as hyponatremia following brain trauma, concussions or MDMA toxicity.